فتوسنتز در گیاهان c3,c4,cam

مقدمه:
فتوستز فرایندی است که طی آن انرژی نورانی خورشید به انرژی شیمیایی تبدیل می شود. که مهمترین پدیده بیولوژیکی روی کره زمین است که منبع انرژی اولیه بشر است. پس فتوسنتز اساس رشد و عملکرد تمام گیا هان زراعی می باشد.
فتوسنتز فقط در گیا هان سازگار از نظر ژنتیکی رخ می دهد.و گاهی متغیر های محیطی نیز ممکن است اثرات محیطی را بپوشاند. از نقطه نظر اصلاح نباتات تلاش برای افزایش عملکرد دانه بدون توجه به مقدارمواد فتوسنتزی قابل دسترس ، به طور کلی شکست خورده است.
فرایند فتوسنتز :
مکان فتوسنتز کلروپلاست ها می باشد.کلروپلاستها ، اجسام رنگدانه داری (کلروفیل) در ستوپلاسم سلولهای برگ و دیگر بافت های سبز می باشند. اختلافات ژنتیکی و محیطی مقادیر نسبی رنگدانه کلروفیل را تغییر داده و برگهایی را تولید می کنند که رنگ سبز تیره تا زرد دارند.
علاوه بر کلروپلاست Co2 , H2o وانرژی نورانی خورشید نیز لازم است تا گلوکز تولید شود. گلوکز به برگها و یا به دانه ها رفته و تغییر شکل می دهد.
کارایی فتو سنتز :
کارایی فتوسنتز در حدود 1% است.یعنی از کل تشعشعات ورودی به یک مزرعه فقط 1% به انرژی شیمیایی تبدیل می شود. رسیدن به کارایی بالا در فتوسنتز امکان پذیر است ولی در شرایط مزرعه بیش از 6% انتظار نمی رود.حد بالای کارایی تا 20% به صورت فرضی محاسبه شده است.
تاریخچه
کربن (واژه لاتین carbo به معنی زغال چوب) در دوران پیشاتاریخ کشف شد و برای مردم باستان که آن را از سوختن مواد آلی در اکسیژن ضعیف تولید میکردند، آشنا بود.(تولیدزغال چوب).مدت طولانی است که [الماس] بهعنوان مادهای زیبا و کمیاب به حساب می آید .
فولرن ،آخرین آلوتروپ شناخته شده کربن در دهه 80 بهعنوان محصولات جانبی آزمایشات پرتو مولکولی کشف شدند. گیاهان CAM روزنه های خود را در شب باز و در روز می بندند. و این خود کارایی مصرف آنها را افزایش می دهد. و تلفات آب به حداقل می رسد.
از آنجا که مسیرورود و خروج CO2 همان روزنه های برگ است. پس اسیمیلاسیون CO2 در شب صورت می گیرد و این همزمان با کربوکسیلاسیون فسفو اینول پیرووات و اگزالواستات (که تبدیل به مالات میشود) اتفاق می افتد. (فسفواینول پیرووات از تجزیه کربوهیدراتهابوسیله مسیر گلیکولیت حاصل می شود) اسید C4 بصورت مالیک اسید در واکوئلهای بزرگ گیاهان CAM تجمع می یابد.
باشروع روز، روزنه ها بسته می شوندو از تلفات آب و جذب بیشتر CO2 جلوگیری می کنند.و مالیک اسید ذخیره شده در واکوئلها مصرف می شود. دکربوکسیلاسیون از طریق عمل آنزیم مالیکNADP برروی مالات یا فسفواینول پیرووات کربوکسی کیناز برروی اگزالواستات صورت می گیرد. CO2 آزاد شده نیز وارد چرخه
C3 PC شده و تبدیل به کربوهیدرات می شود.
بالارفتن CO2 درونی، اکسیژناسیون ریبولوز بی فسفات رامتوقف نموده و به کربوکسیلاسیون کمک می کند.اسید C3 باقیمانده در اثر دکربوکسیلاسیون، ابتدا به تریوز فسفات و سپس به نشاسته یا قند تبدیل می شود.
لازم به ذکر است که طبق مشاهدات به عمل آمده مشخص شده است که کربوکسیلاز در شب فعال و در طی روز غیرفعال بوده در عوض دکربوکسیلاز در طی روز فعال و در شب غیر فعال است.
اولین مرحله در واکنشهای تاریکی فتوسنتز، کربوکسیلاسیون یک پذیرنده 5 کربنی ( ریبولوز 1و5 - بی فسفات ) است که نتیجه آن، تولید یک ماده حدواسط در چرخه PCR است. اولین مرحله واکنش تاریکی را آنزیمی بنام « ریبولوز بی فسفات کربوکسیلاز- اکسیژناز» که مخفف آن رابیسکو ( RUBISCO ) است، کاتالیز میکند.
یکی از خصوصیات جالب رابیسکو اینست که نه تنها میتواند ریبولوز1و5- بی فسفات را کربوکسیله کند، بلکه آن را اکسید نیز میسازد. که این آغازگر فرایند دیگری بنام تنفس نوری است. این بدلیل آنست که کربوکسیلاسیون و اکسیژناسیون هر دو در یک محل فعال از آنزیم اتفاق می افتد. البته باید متذکر شد که میل ترکیبی رابیسکو نسبت به CO2 بسیار بیشتر از O2 است.
یعنی در هوای آزاد نسبت کربوکسیلاسیون به اکسیژناز 3 به 1 است. بجز غلظت CO2، درجه حرارت نیز بر حلالیت نسبی CO2 و O2 تاثیر دارد. افزایش دما باعث افزایش تنفس نوری در مقایسه با فتوسنتز میشود.
البته چرخه دیگری در گیاهان بنام چرخه C2PCO وجود دارد. که فسفو گلیکولات ساخته شده در جریان اکسیژناسیون ریبولوز بی فسفات وارد آن شده و می تواند طی واکنشهای پیاپی که در کلروپلاست، پراکسی زوم و میتوکندری صورت میتواند حدود 75 درصد از کربن را دوباره بازیافت کرده و به چرخه C3PCR بازگرداند.
از آنجا که فتوسنتز و تنفس نوری در خلاف جهت یکدیگر فعالیت میکنند. لذا تنفس نوری باعث اتلاف CO2 در سلولهایی میشود که همزمان از طریق چرخه PCR به تثبیت CO2 مشغولند.رقابت میان واکنشهای کربوکسیلاسیون و اکسیژناسیون در گیاه باعث کاهش کارایی ترمودینامیکی فتوسنتز می شود. بعنوان مثال در هوای معمولی، اکسیژناسیون ریبولوز بی فسفات و وجود چرخه , PCO انرژی لازم برای تثبیت یک مول CO2 را از521 میکروژول به 867 کیلو ژول افزایش میدهد.
* تعدادی از گیاهان فاقد تنفس نوری اند. این وضعیت بخاطر متفاوت بودن خاصیت رابیسکو نیست. بلکه بدلیل وجود مکانیسمی است که CO2 را در محیط عمل رابیسکو تغلیظ میکند.هنگامیکه غلظت CO2 در حد بالایی باشد،واکنش اکسیداسیون متوقف میشود.
- سه مکانیسم برای تغلیظ CO2 در محل کربوکسیلاسیون وجود دارد.
1- پمپ CO2 : که در گیاهان آبی، جلبکها و سیانوباکتری ها دیده شده است. این پمپها با صرف
انرژی حاصل از واکنشهای نوری باعث تجمع کربن غیر آلی بصورت CO2/HCO3- میشوند. که این امر موجب اختلاف بسیار زیاد کربن غیر آلی درون سلول نسبت به بیرون(حتی بیشتر از3 برابر) میشود.این پمپها در سلواهایی که در غلظت بالای CO2 رشد میکنند وجودندارد.
نتیجه متابولیکی افزایش CO2اینستکه مانع ازاکسیژناسیون ریبولوز بی فسفات میشود.و ازاینروباعث توقف تنفس نوری میشود.
2- چرخه اسیمیلاسیون کربن فتوسنتزی C4 (PCA) :
این چرخه هم در گیاهان تک لپه و هم در دو لپه ایهادیده شده ولی بیشتر در گونه هایی از غلات، چغندریان و جگنها دیده میشود.گیاهان این چرخه از نظر آناتومی با سایر گیاهان متفاوتند.
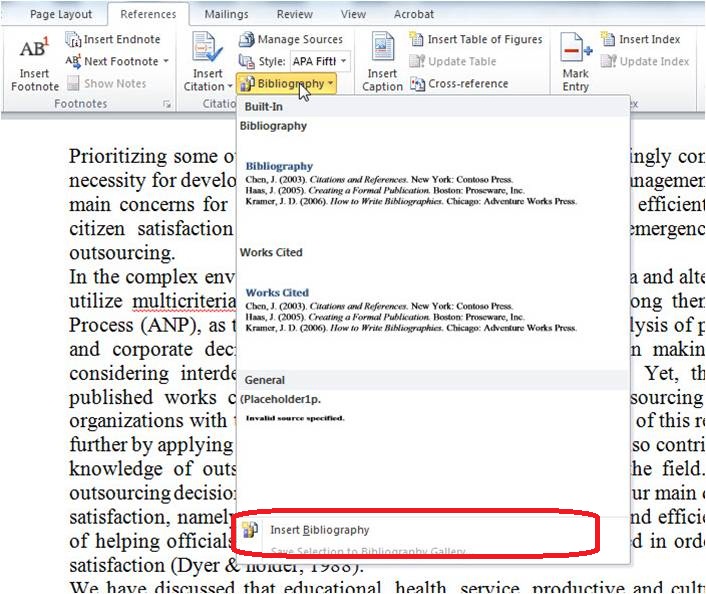
مشاهده ساختمان برگ گیاهان C4، C3و CAM
تقسیم بندی گیاهان براساس نحوه فتوسنتزو محتوای کلروپلاستی وانزیمیو اولینترکیبات حامل از تثبیت co2به دو گروه :
الف) گیاهان C3:مثل جلبک خزه ها وهمه درختان ،اولین ترکیبات حامل از تثبیت co2 :اسید فسفرگلیسریک c2
ب) گیاهان C4: مانند ذرت ، نيشكر، تاج خروس همگی علفی اند اولین ترکیبات حامل از تثبیت
co2:اسید اگزالواستیک
در گياهان مختلف سبزينه دار كه فتوسنتز انجام مي شود و در نتيجه مواد غذايي ساخته مي شود اولين محصول پايدار بدست آمده در آنها متفاوت است . بدين معني كه اولين محصول پايدار در دسته اي از گياهان يك اسيد سه كربني به نام «3- فسفو گليسيريك اسيد» و دسته اي ديگر يك اسيد چهار كربني به نام« دي كربوكسيليك اسيد » ( داراي دو عامل كربوكسيل COOH- ، مثل اسيد اگزالواستيك ،اسيد ماليك ، اسيد آسپارتيك ) مي باشد .
گياهاني كه اولين محصول پايدار حاصل از فتوسنتز آن ها يك اسيد سه كربنه است گياهان C3 و آندسته كه اولين محصول پايدار آن ها چهار كربني است گياهان C4 ناميده مي شوند . گياهان C4 در مقايسه با گياهان C3 از بازدهي فتوسنتزي بيشتري برخوردارند . بر همين پايه علف هاي هرز C4 نسبت به علف هاي هرز C3 قدرت رقابت زيادتري دارند . در تعدادي از گياهان گوشتي فرآيند فتوسنتزي ديگري مشاهده شده كه در شرايط رطوبت كم روزنه ها در شب باز شده و Co2 جذب مي كنند و در روز بسته مي شوند لذا شدت تعرق گياه خيلي كم مي شود . به اين نوع مكانيسم ، متابوليسم كراسولايي (CAM) "Crassulation asid metabolism" مي گويند مانند آگاو ،آناناس ،كاكتوس . كليه ي گياهان (CAM) جزءگياهان گوشتي غير نمكدوست هستند و عموما با محيط هاي خشك سازگارند . برخي از گياهان C3 عبارتند از گندم ،جو ،سلمه ،ترشك ، توق ، تاتوره ،يولاف،بارهنگ و پنيرك .
از گياهان C4 مانند ذرت ، نيشكر ، اويار سلام ، قياق ،سوروف ، پنجه مرغي ،تاج خروس ،خرفه ،سورگوم ،علف شور ،خار خسك .
تیپ C3 که در اکثر گیاهان دیده می شود، به دلیل اینکه CO2 تثبیت شده در ساختار گیاه، در اولین مرحله از واکنشهای تاریکی فتوسنتز (چرخه کالوین) توسط آنزیم روبیسکو منجر به تولید قندی 3 کربنی موسوم 3- فسفو گلیسر آلدهید میشود، به این نام معروف است. این گروه از گیاهان با یک مشکل جدی مواجه هستند.
تنفس نوری
این فرآیند که به دلیل عملکرد دوگانه روبیسکو (کربوکسیلاسیون و اکسیژناسیون) رخ میدهد، منجر به کاهش بازده فتوسنتز در گیاهان میشود. دلیل این هم که این گیاهان بر خلاف گیاهان تیپ C4 قادر به حذف چنین فرآیندی نیستند به ساختار این دو دسته گیاهی برمیگردد: در گیاهان C4 محل انجام مرحله روشنایی فتوسنتز (مراحل مربوط به انتقال الکترون بین فتوسیستم I و II که منجر به رهاسازی اکسیژن به عنوان عامل آغازگر تنفس نوری میشود)
از محل انجام واکنشهای مرحله تاریکی (چرخه کالوین که روبیسکو در این چرخه قرار دارد) جدا است! به طوری که واکنش اول در سلولهای مزوفیلی و واکنش دوم در سلولهای غلاف آوندی رخ میدهد. این امر موجب می شود که اصولا اکسیژن آزاد شده طی واکنشهای روشنایی در مزوفیل به دلیل عدم دسترسی به روبیسکو به عنوان عامل تنفس نوری (که در درون سلولهای غلاف آوندی است)قادر به ایجاد تنفس نوری نباشد .
در طرف مقابل ، در گیاهان C3 چنین تمایز سلولی در انجام دو مرحله فتوسنتزی وجود نداشته و هر دو مرحله در سلول مزوفیلی رخ داده و اکسیژن آزاد شده طی واکنشهای نوری به عنوان سوبسترایی رقابتی با دی اکسید کربن بر روی جایگاه فعال آنزیم روبیسکو به رقابت پرداخته و منجر به وقوع تنفس نوری شود که این امر موجب کاهش بازده فتوسنتز در این گیاهان می شود . اصولا میزان تمایل روبیسکو به دی اکسید کربن خیلی بیشتر از تمایل آن به اکسیژن است. ولی به هر حال همین مقدار تنفس نوری هم میتواند منجر به کاهش بازده تنفس نوری شود.
در گياهان مختلف سبزينه دار كه فتوسنتز انجام مي شود و در نتيجه مواد غذايي ساخته مي شود اولين محصول پايدار بدست آمده در آنها متفاوت است . بدين معني كه اولين محصول پايدار در دسته اي از گياهان يك اسيد سه كربني به نام «3- فسفو گليسيريك اسيد» و دسته اي ديگر يك اسيد چهار كربني به نام« دي كربوكسيليك اسيد » ( داراي دو عامل كربوكسيل COOH- ، مثل اسيد اگزالواستيك ،اسيد ماليك ، اسيد آسپارتيك ) مي باشد . گياهاني كه اولين محصول پايدار حاصل از فتوسنتز آن ها يك اسيد سه كربنه است گياهان C3 و آندسته كه اولين محصول پايدار آن ها چهار كربني است گياهان C4 ناميده مي شوند . گياهان C4 در مقايسه با گياهان C3 از بازدهي فتوسنتزي بيشتري برخوردارند . بر همين پايه علف هاي هرز C4 نسبت به علف هاي هرز C3 قدرت رقابت زيادتري دارند . در تعدادي از گياهان گوشتي فرآيند فتوسنتزي ديگري مشاهده شده كه در شرايط رطوبت كم روزنه ها در شب باز شده و Co2 جذب مي كنند و در روز بسته مي شوند لذا شدت تعرق گياه خيلي كم مي شود . به اين نوع مكانيسم ، متابوليسم كراسولايي (CAM) "Crassulation asid metabolism" مي گويند مانند آگاو ،آناناس ،كاكتوس . كليه ي گياهان (CAM) جزءگياهان گوشتي غير نمكدوست هستند و عموما با محيط هاي خشك سازگارند .
برخي از گياهان C3 عبارتند از گندم ،جو ،سلمه ،ترشك ، توق ، تاتوره ،يولاف ،بارهنگ و پنيرك .
مقایسه فتوسنتز در گیاهان C3 ، C4 و CAM
|
C3 |
C4 |
CAM |
|
آنزیم تثبیت کننده CO2 |
روبیسکو |
غیراز روبیسکو |
غیر از روبیسکو |
|
محل تثبیت CO2 |
بستره ي کلروپلاست |
سلول های میانبرگ |
واکوئل سلول |
|
مرحله تثبیت CO2 |
چرخه کالوین |
قبل از چرخه کالوین |
قبل از چرخه کالوین |
|
زمان باز شدن روزنه های هوایی |
روز |
روز |
شب |
|
زمان تثبیت CO2 |
روز |
روز |
شب |
|
زمان چرخه کالوین |
روز |
روز |
روز |
|
زمان تولید قند در فتوسنتز |
روز |
روز |
روز |
|
مرحله ساخته شدن قند در فتوسنتز |
چرخه کالوین |
چرخه کالوین |
چرخه کالوین |
|
محل ساخته شدن قند در فتوسنتز |
تمام سلول های فتوسنتز کننده |
سلول های غلاف آوندی |
تمام سلول های فتوسنتزکننده |
|
اولین ترکیب پایدار حاصل از تثبیت CO2 |
اسید سه کربنی |
اسید چهار کربنی |
اسید چهارکربنی |
گیاهان C3 و C4 :
زمانی که اولین محصول فتوسنتزی در گیاه ،یک ترکیب 3 کربنه (C3) باشد از مسیر کالوین – بنسون و اگر 4 کربنه (C4) باشد از طریق هچ – اسلک است .که برای اصلاع گیاهان C4 به کار برده می شود. و انحراف از نوع فتوسنتز C4 را متابولیسم اسید کراسولاسه یا CAM می نامند که با مناطق خشک سازگاری دارد.
گیاهان C4 (ذرت ، سورگوم ، ارزن) در شرایط گرم و نور زیاد کارایی بیشتری از گیاهان C3 (غلات) دارند . در مجموع کارایی آنها (C4) حدود 40 % است.
تمام گیاهان به انرژی نیاز دارند و این انرژی از طریق تنفس به دست می آید :
2 نوع تنفس وجود دارد :
1- مستقل از نور : قند و کربو هیدرات تولید شده در فتوسنتز را به CO2 و H2O تبدیل می کند. از نظر زراعی تنفس نشان دهنده تلفات است و باید به حداقل برسد که درست نیست زیرا تنفس لازم است تا انرژی برای رشد و بقای گیاه فراهم شود و تنفس بیش از نیاز برای تولید عملکرد زاید است . پس اهمیت سرعت اسیمیلاسیون خالص بیش از سرعت فتوسنتز و تنفس است.
2 – تنفسی که در حضور نور و در گیاهان C3 مشاهده می شود و ظاهرا در گیاهان C4 وجود ندارد.
اختلافات گیاهان C3 و C4 :
1 – گیاهان C4 نسبت به C3 به نورهای با شدت بالا واکنش نشان می دهند.(2 برابر گیاهان C3)
2 – گیاهان C4 با کارایی بیشتری از CO2 استفاده می کنند و در شدت نور ثابت و نسبتا زیاد قادرند CO2 اطراف را به ppm 10 – 0 کاهش دهند. که این نقطه جبران است . در حالی که در C3 برابر
ppm 150 -50 co2 است . نقطه جبرانی پایین نشان دهنده کارایی زیاد فتوسنتز است .
3- گیاهان C4 سرعت اسیمیلاسیون خالص بالاتری از C3 دارند که به خاطر عدم وجود تنفس نوری است .
4 – درجه حرارت بر روی C4 تاثیر مطلوب ، و بر روی گیاهان C3 تاثیر نا مطلوبی دارد . تا دمای 0 تا 35 درجه سانتیگراد سرعت تنفس به ازای هر 10 درجه سانتیگراد افزایش حدود 2 تا 4 برابر افزایش می یابد. بسیاری از گیاهان C3 در محدوده دمایی 25 تا 35 درجه سانتیگراد تولیدی ندارند ولی گیاهان C4 افزایش نولید دارند.
5 – در گیاهان C4 کارایی مصرف آب بیش از گیاهان C3 است . میانگین ماده خشک تولید شده برای هر 1000 گرم آب مصرفی ، 29/3 گرم برای C4 و 54/1 گرم برای C3 می باشد.
6 – سرعت انتقال با سرعت فتوسنتز همبستگی دارد زیرا انتقال آهسته می تواند عملکرد را محدود کند . در برگهای گیاهان C4 انتقال 2 برابرسریعتر از برگهای C3 است.
7 – حداکثر رشد گیاهان C4 بیشتر از گیاهان C3 است (به جز چند استثناء). وقتی طول فصل رشد متوسط در نظر گرفته می شود سرعت رشد C3 برابر 13 گرم در متر مربع در روز بوده و برای گیاهان C4 برابر 22 گرم در متر مربع در روز می باشد. با محصور کردن گیاهان C3 و C4 در ظرف در بسته دارای نور اختلاف در تنفس نوری را می توان نشان داد . چون C4 نقطه جبرانی پایین تری از گیاهان C3 دارند زمانی که گیاهان C4 از CO2 مصرف می کنند گیاهان C3 از بین می روند.
فتوسنتز و عملکرد غلات :
چون کربن جزء اصلی ماده خشک گیاه است ، پس منطقی است که افزایش سرعت اسیمیلاسیون دی اکسید کربن باعث افزایش عملکرد شود. از حدود سالهای 1960 سرعت اسیمیلاسیون Co2 نقطه شروعی برای توجیه بهبود عملکرد گیاهان زراعی بوده است و درک اینکه چرا افزایش سرعت فتوسنتز برگ ممکن است عملکرد را افزایش ندهد اساس درک فیزیولوژی در رابطه با رشد و نمو آن ها می باشد.
تمام مواد فتوسنتزی تولید شده به سمت عملکرد اقتصادی یعنی دانه هدایت نمی شود . بخشی به ریشه ها ، برگ ها ، ساقه ها و وظایف حیاتی گیاه مصرف می شود . لذا اندازه گیری سرعت فتوسنتز یک برگ از یک گیاه منفرد، نمی تواند نشان دهنده کل سرعت فتوسنتز آن گیاه با جمعیت گیاهی باشد.
زلیچ دریافت که اندازه گیری فتوسنتز گمراه کننده است و دریافت که عملکرد گیاه زراعی رابطه نزدیکی با اسیمیلاسیون فتوسنتزی خالص Co2 دارد که در طول فصل محاسبه شده باشد .و همچنین آزمایشات ، بر روی غنی سازی Co2 حاکی از آن است که افزایش فتوسنتز خالص عملکرد را افزایش می دهد.
اختلاف در کارایی فتوسنتز :
برخی گونه های گیاهی زراعی سرعت فتوسنتزی متفاوتی را نشان می دهند . بعنوان مثال سورگوم و ذرت ظرفیت بهره برداری انرژی نورانی خورشید را با سرعت تقریبا 50 تا 66 % بیشتر از توتون دارند . که بخش عمده ای از این اختلاف را می توان با اختلافات در تنفس توضیح داد.
نمو گیاه و فتو سنتز :
آندوسپرم بذر غلات زمانی تا مین کننده غذا است که بذر شروع به جوانه زنی و کلئوپتیل خارج و اولین برگ شروع به فتوسنتز نمایید. در شرایط گرم و مرطوب طی 4 تا 5 روز ، و در شرایط سرد و خنک تا 2 هفته طول می کشد. البته عمق کاشت و وجود سله در فواصل بین بذر کاری و سبز شدن نیز تاثیر دارد. بعد از سبز شدن و رشد سریع سطح فتوسنتزی کود را به صورت نواری در 2 طرف بذر و کمی پایین تر قرار می دهند.
اگربذر در تماس مستقیم با کود قرار بگیرد ، ممکن است سبب ضایعات اسمزی شود . مرحله رشد رویشی (جوانه زنی – گلدهی) ، که طی آن میانگره ها و تعداد پنجه ها ، تعداد و اندازه سلول ها ، طول میانگره ها تحت تاثیر طول مدت مرحله رویشی که خود نیز تحت تاثیر درجه حرارت و طول روز است .افزایش قطر ساقه ها از طریق بزرگ شدن سلول ها زمانی رخ می دهد که ساختمان های ضروری شکل گرفته اند.
مرحله رسیدگی از زمان تلقیح گل تا بلوغ بذر می باشد.این مرحله برای توسعه دانه بحرانی است و مصرف آب ، سطح برگ برای جذب موثر نور خورشید ، یا اضافه کردن مواد غذایی ممکن است فتوسنتز را برای توسعه دانه افزایش دهد ،بدون آنکه رشد رویشی را تحریک کند.
عملکرد وزن خشک نهایی دانه حاصل سرعت پر شدن دانه و طول مدت پر شدن دانه می باشد. و زمانی که در وزن خشک دانه افزایشی صورت نمی گیرد محصول به بلوغ فیزیولوژیکی خود رسیده است . که با نمونه برداری از دانه و خوشه می توان توقف افزایش وزن خشک و مرحله برداشت را تعیین نمود.
روابط مخزن – منبع :
برای تولید موفقیت آمیز غلات باید به بذر کاری در زمان مناسب و ظرفیت فتوسنتزی مناسب برای تولید دانه توجه شود.
در جو، منبع و مخزن به نظر نمی رسد که هیچ کدام محدودیتی برای عملکرد دانه داشته باشند و اثرات متقابل فید بک ها نشان می دهد که سرعت فتوسنتز ممکن است برای نیاز دانه تعدیل شود. در غلات اندازه مخزن به تعداد دانه در سنبلچه و حجم دانه و وزن آن بستگی دارد . مطالعه ای در گندم و برنج نشان می دهد که ظرفیت ذخیره ممکن است محدودیت اصلی برای عملکرد دانه باشد